
![]()
|
|
|
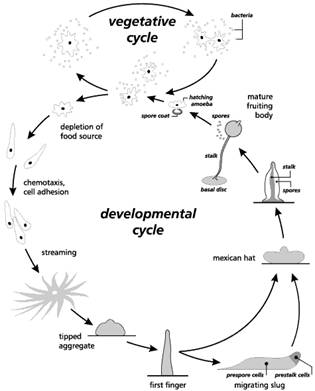
LAB 1 CELL SIGNALING IN DICTYOSTELIUM I. Introduction We will be introducing a number of model organisms throughout the semester that can be used to help in our understanding of development. Dictyostelium is a fascinating organism that offers many advantages for studies of development. Dictyostelium is also fairly easy to propagate, and the complete life cycle can be induced in the laboratory. The cellular slime molds were formerly considered to be 'lower fungi.' Although they superficially resemble fungi in some respects, they are currently included in the Kingdom Protista. Individual cells resemble small amoebae and move and feed in an amoeboid manner, thus they are called 'myxamoebae' (to distinguish them from true amoebae). Dictyostelium was first discovered in 1935 in a forest in North Carolina and has since been found, along with similar genera, in many such environments around the world. The myxamoebae feed on bacteria in decaying vegetation, and reproduce by binary fission as do all amoeba. Cellular slime molds undergo a unique series of developmental events that occurs when food sources become scarce. The individual cells begin to associate, forming streams of migrating cells which merge in an aggregate consisting of up to 100,000 cells. The multicellular aggregate secretes material that forms a sheath around the entire structure, which forms a tower of cells. In the absence of light the tower topples over and starts to migrate as a single organism, called a pseudoplasmodium, slug, or grex. The slug resembles a blob of petroleum jelly, leaving a trail of slime as it migrates. Following migration (or immediately upon aggregating if there is plenty of light), the slug contracts and the anterior tip begins to rise to form a fruiting body. As the fruiting body forms, the cells differentiate into a base, stalk, and apical mass containing encapsulated spores. The spores can tolerate a wider range of environmental conditions than myxamoebae, and germinate following dispersal, renewing the cycle.
Here are some notes on its lifecycle: 1. Vegetative Stage – This stage starts with the hatching of the myxamoebae from spores. Under favorable conditions of moisture and warmth each spore releases a single myxamoeba (also called amoeba) which crawl on the agar substrate (in our experiments) and feed on bacteria. The myxamoeba are attracted to the bacteria by the folic acid that the bacteria secrete. 2. Aggregation - When food supply is limited the amoeba stop moving for 4-8 hours and it looks like nothing is happening. In fact, the amoeba are activating over 100 new genes that will allow for individual cells to find one another and to stick together when they do. Examples of some of the key genes and products include: Adenylate Cyclase that makes cAMP cAMP receptor proteins placed on the cell membrane Phosphodiesterase an enzyme complex the breaks down cAMP Phosphodiesterase inhibitor which blocks the action of phosphodiesterase Cell surface glycoproteins that facilitate cell adhesion After this period individual cells start to secrete cAMP which acts as a chemotatic agent and cells start to move toward the area of highest concentration of cAMP. There is a regular cycle of secretion of cAMP and movement that lasts for about 60 seconds, then the cells wait for about 5 minutes, then there is another pulse. When cells bump into each other they stick via their new cell surface glycoproteins. 3. Pseudoplasmodium - The previous step of aggregration takes 8-12 hours from the start of starvation. The aggregate of cells forms a slug, grex or pseudoplasmodium made up of anywhere from a few hundred to over 100,000 cells. These guys move and have anterior end which is attracted to light, migher temperatures and higher humidity. Their size is variable, but they are normally 2-4 mm in length and not easy see, but their trail made up of a cellulose sheath is easy to visualize. 4. Culmination - The slug transforms into a structure called a fruiting body by first settling down on its posterior end and looks like a Mexican hat. Cells reorganize themselves prespore and prestalk cells. Over the next 8-10 hours a mature fruiting body forms which is normally 1-2 mm high. Dictyostelium discoideum is extremely resistant to damage. You can hit it with a hammer and find that you’ve merely asked an interesting question: “What happens when the organism is disaggregated?” Also, the organism’s simplicity invites us to examine complex principles in the context of a minimum number of different types of cells involved in the process. Developmental biologists have recognized these advantages for years and have used Dictyostelium to ask questions about mechanisms of cell movement and chemotaxis, about the biochemistry of differentiation, and about the genes involved in differentiation and pattern formation. Though genetic manipulations are beyond the scope of this laboratory, many other realms are open to you. Use your imagination. Design simple experiments that will yield sophisticated answers. Keep in mind that a chemical usually has more than one effect. And enjoy this experimental playground.
II. Procedures: For this lab you will be designing and performing your own experiments. What follows are suggestions of techniques you can use. When you start and experiment, state clearly in the “Experimental Setup” section of your laboratory notebook the question or questions you are asking. This is absolutely essential. You need to know what you are asking before you can know what the answers are. In addition to your observations of normal development, it is suggested that you do several experiments that are variations on the four themes below: 1. Labeling with vital dyes You can mark individual organisms or regions of organisms with vital dyes. A vital dye is one that is not toxic to cells when used in low concentrations. It is toxic at high concentrations, however; the rule of thumb is that a vital dye should not be used at concentrations higher than 0.1%. This is considered a stock solution. The solution must be sterilized before use. This can be done by filtering it through a 0.22-mm porosity filter into a sterile test tube. When choosing a dye, you should always be aware of what elements in the cell the dye stains. Often valuable biochemical information can be gained by noticing the pattern of staining. Useful dyes include Nile blue sulfate, neutral red, methylene blue, and toluidine blue (Table 4.1). You can use a vital dye to stain any of the stages in the life cycle of the slime mold. You could stain one group of amoebae and follow them through the rest of the life cycle. You could stain a slug and look for differential staining between prestalk and prespore cells. Move the structure you are staining to an unused quadrant of your culture dish (unless you are staining aggregating amoebae, which should be left in place so as not to disturb their configuration—unless a disturbance is part of the experiment design). Place a drop of dye directly on the cells you are staining. Excess dye will sink into the agar and doesn’t need to be removed. Record which stain you used. Diagram the staining pattern. If you used a metachromatic dye, note regions of primary and secondary colors. What conclusions about biochemical differences can you draw from the differences you see? Observe the organism throughout the rest of its life cycle, noting any changes in color patterns.
2. Transecting, grafting, and disaggregating pseudoplasmodia Pseudoplasmodia (slugs) can be transected (cut) into halves, thirds, and so on. They also can be disaggregated into individual cells. A number of interesting questions can be asked in this way. For example, are the fates of the cells of a slug irrevocably set? If a slug is cut in half, what will happen? Can a half-slug survive? Can it differentiate? If so, what will it form? Will the anterior half form only stalk, or will some of the cells alter their fate to become spore cells, thus forming an entire fruiting body? If so, is the fruiting body that results normal in its proportions? Do the results vary depending upon how much time the slug wanders before it settles down to differentiate? What happens if you cut off just the tip of a slug? What do your results say about the flexibility of cells whose fates have already been determined? The more questions you ask of a single experiment, the more acute your observations will be, and the more data you will collect from each experiment. Cool the slug down first for 10 minutes by placing plate in frig or on ice. To divide a slug into two or more pieces, first move the slug to an unused quadrant of your culture dish. This can be done by lifting the slug on the flat side of a microknife, or even by lifting out a section of agar along with the slug so as not to risk hurting the slug. You will have an easier time cutting the slug if it is thoroughly cooled first. Place your dish in the refrigerator for 10 minutes, or place it on crushed ice held in a larger dish. Use a microknife to cut the slug. Cuts can be made through the slug into the agar below. The knife is ten drawn through the agar without touching the cells. Always make careful diagrams of the position of your cuts. Devise a scheme that will allow you to distinguish one piece from another. For example, each piece could be stained with a different vital dye. Cutting the slug often results in bursting the cellulose sheath that enclosed the cells, and instead of two discrete pieces you have a puddle of disaggregated slug. This is fine—don’t despair. Your experiment has simply changed to being a disaggregation experiment. What will happen when you disaggregate a slug? Will the cells reaggregate? If so, will they reaggregate immediately, or will there be a lag period prior to reaggregation? What does this mean at the biochemical level? Will the disaggregated cells behave differently if you give them food (E. coli)? Can you force the life cycle to go backward? Just how flexible is a cell that has already become differentiated as a prespore or prestalk cell?
Grafting one piece of a slug onto another is rather more difficult than a simple cut, but can be used to ask more complex questions. What would happen if two anterior ends were grafted together? Would there be a battle for anterior dominance? In which direction would the resulting slug move? What would it develop into? The difficult part of grafting is keeping the two pieces together until they have annealed (joined together). It will help if you cool the slugs before cutting. It will also help if the slugs to be cut are close together initially, which diminishes the distance each piece must be moved. Choose the pieces you want to anneal, transfer one to the other by lifting it on the flat side of a microknife or other suitable instrument, and gently push the two pieces together. It will help to keep the dish cool during the first few hours of annealing. You could also dig a shallow depression in the agar where the pieces could sit. The shape of the depression should be such that it helps to force the pieces together. Use diagrams to indicate exactly the regions that were used in the graft. 3. Disruption of cyclic AMP levels Altering cAMP levels will undoubtedly have profound effects on the organism. It is important to know that exogenous (extracellular) cAMP will not cross cell membranes to enter a cell and raise intracellular cAMP levels. There are various chemicals that you could use to disrupt extracellular levels of cAMP. You could use cAMP itself, soaked into an agar cube or flooding an entire area of agar. The concentration of cAMP you use is your choice, but you should at least know some effective ballpark figures. Aggregating amoebae can respond to cAMP levels as low as 0.01 mg/ml. Levels used in chemotaxis experiments are often much higher than this, ranging from 0.05 to 1.2 mg/ml. At 1.2 mg/ml, amoebae fail to continue development. Since cAMP may be something your laboratory cannot supply, there are other sources readily available. The tip of the migrating slug, for example, secretes cAMP and can be removed and placed among amoebae. Also, human urine has relatively high levels of cAMP (approximately 0.53-0.75 mg/ml), well within the range that amoebae respond to. Urine, of course, is a complex substance, and this must be kept in mind when you are drawing your conclusions. For example, urine also contains cGMP (approximately 0.09-0.23 mg/ml), which acts to block the cells’ phosphodiesterase. (This should actually augment the rise in extracellular cAMP levels.) If you use urine, measure its pH (on average, this will be pH 6.3, but normal ranges are from pH 4.5 to pH 8.0), and be aware that it contains some ammonia (0.07-0.13% on average). Basic pH and ammonia are known to favor spore formation and decrease stalk formation. Another method of altering cAMP levels is indirectly, by stimulating or inhibiting the enzymes involves in making cAMP. Dictyostelium’s phosphodiesterase, the enzyme that breaks down cAMP, is secreted at low levels during the amoeboid stage, and it is found at much higher levels, inserted in the membrane just prior to aggregation. During the amoeboid stage, the extracellular phosphodiesterase probably is useful in breaking down the cAMP secreted by bacteria. This would prevent the amoebae from responding prematurely to cAMP. During aggregation, the membrane phosphodiesterese cleans off cAMP binding sites, getting them ready for each new pulse of cAMP. What do you think would happen if you stimulated or inhibited phosphodiesterase during aggreagation? In higher organisms, caffeine or theophylline could be used to inhibit phosphodiesterase, thereby raising cAMP levels, but the phosphodiesterase of Dictyostelium is relatively unresponsive to these substances. Caffeine at a concentration of 10 mM (0.194 mg/ml) does not inhibit it, and theophylline will decease its activity by 15% when present at a concentration of 10 mM (1.8 mg/ml). In fact, caffeine’s effect on Dictyostelium is just the opposite of what you might expect. It actually indirectly increases levels of phosphodiesterase activity by inhibiting adenylyl cyclase. Intracellular cAMP levels drop, and this, in turn, may raise phosphodiesterase activity. 5 mM caffeine (0.97 mg/ml) can double the activity levels of phosphodiesterase in Dictyostelium amoebae. If you choose to use caffeine or theophylline, you can use the purified form, or a cheaper source—that cup of coffee or tea you’re not allowed to drink in the laboratory. Drinking strengths of coffee contain 0.5 to 1 mg/ml of caffeine, and a cup of tea has about 0.3 mg/ml of caffeine and 0.04 mg/ml of theophylline; these levels are well within the range that should affect amoebae. You may find, as J. T. Bonner did, that as little as 0.025 mg/ml of theophylline can delay aggregation. How might you explain this at the biochemical level? What do you think the effect of caffeine or theophylline would be on a migrating slug or Mexican hat? If you use coffee or tea to ask these questions.
4. Behavioral studies Prior to starvation, Dictyostelium amoebae show chemotactic behavior toward the folic acid (one of the B vitamins) that bacteria secrete. You could test the various aspects of this behavior. What concentration of folic acid is effective? (A range between 10-6 M and 10-2 M are ballpark concentrations to try: folic acid can be bought as a vitamin supplement, a 400 mcg pill dissolved in 10 ml distilled H2O makes approximately a 10-4 M solution.) are cells of a disaggregated slug attracted to folic acid? Dictyostelium slugs show a number of behaviors. For example, they migrate toward light, higher temperature, higher humidity, low ionic concentrations, and the acidic side of a pH gradient. The cue for settling down into a Mexican hat may be the breaking of the surface film of water by the anterior tip of the slug, which it would do under normal circumstances if it were moving upward, toward light. The presence of ammonia, which is copiously produced by the migrating slug, inhibits culmination. You can design experiments that will test various parameters of behavior. For example, will slugs settle into Mexican hats in the absence of light? Are they more sensitive to certain wavelengths of light than others? If you expose a slug to ammonia will you prevent culmination? And if so, for how long? (10-100 mM ammonium hydroxide titrated to pH 7.5 with concentrated phosphoric acid is a concentration range you could use; household ammonia is approximately 3-4—diluting it to mM concentration is something you could try.) What salts encircling an aggregate or slug prevent migration, and why do you think migration is inhibited? (It is presently not known why.) A number of questions can be asked. The trick is to design a setup that will do the asking. A box with a small hole at one end provides a good chamber for testing movement toward light, and different colors of cellophane can be used over the hole to test different wavelengths of light. You will have to observe your cultures over several days to collect your data. Dictyostelium fruiting bodies are surprisingly reactive. They move in response to air currents, for example. And if a dissecting needle is placed near the spore mass, the fruiting body may bend in response to the needle. You can experiment with a number of stimuli (sound, air currents, heat) or substances of different composition (wood, plastic, different metals) to try to determine what specifically the fruiting body responds to. You can also try to determine the stimuli that lead to the release of spores. Is it a response to physical touch, to water? Since these aspects of Dictyostelium biology have been largely ignored by researchers, your results can add new information to our knowledge of this organism. III. QUESTIONS Watch your cultures carefully throughout the rest of the laboratory period—a lot can happen in only a few hours. Also, check your cultures throughout the week as time allows. Keep careful records of any changes. Always use diagrams to document what you see. A great deal of data will be tied up in these diagrams, so make them accurate and proportional. You may run your experiments as long as you wish, or your energy allows. You do not have to terminate them before the next laboratory period. The only constraint is that they be completed before a report on them is due. Have fun and be imaginative.
VI. Materials: Dissecting scopes with substage mirrors and high-intensity lights to be able to visualize slime molds 20 plates of nutrient agar with growing E. coli and Dictyostelium (see culture procedures below) Watchmakers forceps Alcohol lamps or flame source Microscope slides, clear finger nail polish Number 00 or 000 insect pins Old-style thin double-edged razor blades for making micro tools Cotton swabs with long wooden handles (for making micro-tools) Sharpies, Protective eye glasses, leather work gloves for cutting razor blades Heavy duty scissors or small tin snips for cutting razor blades into small pieces Little paraffin pellets for making tools Nile blue sulfate vital dye solution Neutral red vital dye solution Methylene blue vital dye solution Toluidine blue vital dye solution 50 ml of 100 mM caffeine 50 ml of 100 mM theophylline various sizes of cover slips 70% EtOH in squirt bottles Pasture pipettes and bulbs cAMP (if cost allows) 000 insect pins, dental wax and wooden q-tips for making microknives Ph meter with standards 250 ml beakers 10-100 mM ammonium hydroxide titrated to pH 7.5 with concentrated phosphoric acid Sterile distilled water Eppendorf pippetors with fine tips folic acid - (A range between 10-6 M and 10-2 M are ballpark concentrations students will make up: folic acid can be bought as a vitamin supplement, a 400 mcg pill dissolved in 10 ml distilled H2O makes approximately a 10-4 M solution.)
A note to the instructor: Preparing cultures The most economical way of preparing cultures for class is to make subcultures from stock tubes bought from a biological supply house. In this way, just a few tubes of D. discoideum and E. coli can supply enough material for even a hundred students. Timing is the critical factor. If the class is small, making subcultures 5 days in advance of the class and keeping them at room temperature should work well. If the class is large, it is best to make subcultures 13 days prior to class. Use these to seed further cultures for the individual students 5 days in advance of class.
The following is a conventional method for culturing slime molds. Petri dishes may already have been prepared for you with nutrient agar, a culture medium described below. A divided petri dish with four wells of nutrient agar makes an excellent set of experiment chambers. If your petri dish is not divided, you can make barriers with sterilized (use70% alcohol or boiling water) coverslips pushed into the agar. The agar should have a moist but not excessively wet surface. Use sterile distilled water to wet the surface if needed. This culture medium will provide nutrient for Escherichia coli, the bacterium upon which the slime mold amoebae will feed. It is a substrate that is perfectly adequate for all stages of the slime mold’s life cycle. Wherever the amoebae clean out an area of bacteria, they will aggregate to form slugs, and eventually there will be a rich population of slugs and fruiting bodies in the dish. Nutrient agar + E. coli seem to work best for slime mold growth (Devlin, 2007) Sterilize a bacteriological loop in a flame by heating it until it is red-hot. Allow it to cool, then immediately use it to transfer E. coli to the first well or small region of your agar plate. This is done by scraping some bacteria from the surface of the stock tube onto the loop and then lightly moving the loop back and forth over the surface of the agar in your plate. This will create streaks of bacteria. (A suspension of bacteria in sterile distilled water also can be used and smoothed over the surface of the agar with a bent glass rod. This will give an even lawn of bacterial rather than streaks.) Resterilize the bacteriological loop and use it in the same manner as above to transfer mature slime mold fruiting bodies and spores to the same well or small region of the petri dish where the bacteria have been placed. Label the dish with your name and date, and place the dish in a protected area of the lab. The optimum temperature range for growth of the slime mold is 22-24oC (room temperature) and optimum relative humidity is 70-75%. The spores will germinate, releasing myxamoebae that will increase by mitosis and feed on the E. coli. This will continue as long as the supply of E. coli remains. Check on your dish as often as time allows during the week, keeping a precise record of the development you observe. Diagram what you see, note variations among different groups: What are the sizes of the aggregating populations that you see? What sizes of slugs do you observe? What different morphologies do you see among fruiting bodies? By the next week, all states of development should be available to you in your culture dish, since this method does not produce synchronized development. You will do the rest of this lab at that time. If the laboratory instructors have already prepared cultures for you, you will be starting your laboratory exercise at this point. When you receive your dish, answer the questions stated in the paragraph above. Make as many observations as you can about the organism before proceeding to the experimental exercises. Examine different stages under the dissecting scope. Put various stages on a slide, and examine them under the compound microscope. Put a spore mass on a slide, and use a needle to burst it open. Look at the spores under high power; there will be about 70,000 in one spore mass. Each spore is ellipsoid and about 5 mm long. Make diagrams of what you see.
V. References
Tyler, M. S. 2000. Developmental Biology, A guide for experimental study. Sinauer Associates, Second Edition. – Much of this lab is adapted from Mary Tyler’s lab on Dictyostelium Tyler, M. S. and R. N. Kozlowski. 2000. Vade Mecum: An Interactive Guide to Developmental Biology. Sinauer Associates, Sunderland, MA. “Slime Mold.” This chapter of the CD shows, in movies, time-lapse photography, and still pictures, each of the life stages of Dictyostelium and techniques used to set up cultures of the organism. Gilbert, S. F. 2000. Developmental Biology, 6th Ed. Sinauer Associates, Sunderland, MA, Chapter 2. Within this chapter is an excellent, concise summary of the life cycle, biochemistry, and genetics of Dictyostelium. Fink, R. (ed.). 1991. A Dozen Eggs: Time-Lapse Microscopy of Normal Development. Sinauer Associates, Sunderland, MA. Sequence 2 is an excellent abridged version of one of John Tyler Bonner’s films on Dictyostelium. http://www.ruf.rice.edu/~bioslabs/studies/invertebrates/dicty.html http://www.dictyostelium.com/devcyc.gif
| ||||||||||||||||
{kind=link}