![]()
|
|
|
LAB 2 - 3
DROSOPHILA - HEAT SHOCK PROTEINS, CHROMOSOMAL PUFFS AND IMAGINAL DISKS I. Introduction As you know from your study of genetics, a great wealth of genetic information is already available for Drosophila. It is a remarkable model organism for both studies of transmission inheritance, embryonic development and control of gene expression. Aside from its genetics and short life cycle, the fruit fly has some unusual biology, such as polytene chromosomes and imaginal discs, which developmental researchers have put to good use. In this exercise, we will use Drosophila polytene chromosomes and imaginal discs to demonstrate how cells and genes are able to respond to their environment. Or specifically how increasing the temperature can cause the activation of banks of genes that code for heat shock proteins, whose function is to stabilize the 3D structure of critical enzyme systems. We will start by learning how to dissect salivary glands from third instar larvae. We will then prepare polytene chromosomes, identify them and compare the puffing pattern of heat-shocked larvae to that of normal larvae. A. Early Development The oval-shaped egg of a Drosophila is covered with a tough outer layer called the chorion. Sperm enter through the micropyle at the anterior end. Following fertilization, the zygote nucleus undergoes rapid mitotic divisions to produce about 1000 nuclei that localize at the periphery of the cell. This syncytial blastoderm becomes cellularized into distinct cells by forming a cell membrane around each nucleus. The life cycle of this holometabolous insect takes about two weeks to complete. The egg, laid in rotting fruit, hatches into a first instar larva within one day. The larva molts two times to form a third instar larva. The grub or maggot feeds on the rotting fruit. The larva crawls away from the food and onto a dry surface, where it forms a pupa. After about four days, it ecloses (hatches) into an adult fly. The fly is ready to lay eggs within one day of hatching. In a laboratory situation, the life cycle can be speeded up or slowed down by adjusting the temperature, within reason.
B. Chromosomes The regular chromosome complement of Drosophila melanogaster consists of three pairs of autosomes and a pair of sex chromosomes. The stained appearance of mitotic chromosomes in most cells is as follows: Two of the autosome pairs are large and V-shaped (chromosome 2 and 3) whereas the third pair are small dots (chromosome 4). The X chromosome is long and rod-shaped, and a male Y chromosome is J-shaped. In the salivary glands, however, the chromosomes do not appear this way at all. The glands have evolved cells of tremendous size that are controlled by proportionately enlarged nuclei. In the process of enlargement, the chromosomes in each nucleus duplicate themselves many times over, but in such a way that each duplicate is lined up parallel to all other duplicates of the same chromosome, (polyteny). A single salivary chromosome may thus consist of about a thousand copies lined up together. The "banding" of the salivary chromosome actually represents small density differences along the chromosome that are amplified by the fact that the duplicated chromosomes lie in register. When a particular gene is being transcribed, its corresponding band assumes a puffed appearance in which diffuse material extends away from the axis of the chromosome. It is of interest that the same bands are always puffed at a given developmental stage of the larvae and that different bands are puffed at different developmental stages. Their chromosomes can be viewed at the link below: http://www.ncbi.nlm.nih.gov/projects/mapview/map_search.cgi?taxid=7227&query= These eight photomicrographs (courtesy of Dr. Michael Ashburner, University of Cambridge) show the changes in the puffing pattern of equivalent segments of chromosome 3 in Drosophila melanogaster over the course of some 20 hours of normal development. Note that during this period, when the larvae were preparing to pupate, certain puffs formed, regressed, and formed again. However, the order in which they did often differed. For example, in the larva, band 62E becomes active before 63E (c, d, and e), but when pupation begins, the reverse is true (g, h).
During the normal development of the larval salivary gland of Drosophila, considerable changes occur in the patterns of puffing activity. These can be seen as changes in the puffs of the gland's polytene chromosomes, and occur as a consequence of changes in the concentration of the insect's growth and molting hormone, ecdysone. In addition to the changes in gene activity in normal development, there are changes in the activities of a set of genes that occur as a direct consequence of subjecting animals to a wide variety of experimental insults, for example, a brief heat shock. The discovery of the induction of a unique set of puffs by heat shock has led the way to an analysis of gene function and structure in Drosophila that is, so far, unique. If Drosophila are subjected to a brief heat shock (40 minutes at 37EC, the normal culture temperature being 25EC) puffs are induced at specific sites. There are nine inducible puffs in Drosophila melanogaster at bands 33B, 64C, 64F, 67B, 70A, 87A, 87C, 93D, and 95D. Additionally, all other puffs active at the time the temperature shock begins, regress. A second dramatic heat shock response is evident in the pattern of protein synthesis. The synthesis of a small number of heat shock polypeptides (HSPs) is induced and the synthesis of most other proteins ceases; the protein synthesizing machinery is therefore somehow modified so that only HSPs are translated. The cells must have a coordinated response to protect themselves in the event of excess heat; the same responses are also seen as the result of a number of other unrelated environmental insults such as Azide treatment, hydrogen peroxide, valinomycin treatment, or oxygen deprivation. This coordinated response, in which whole batteries of genes are shut down and others are turned on, is thought to be analogous to developmental switching mechanisms such as going from blastula to gastrula or determining dorsal/ventral axes. This coordinated response, exemplified by the heat shock phenomenon, can be thought of as a general mechanism used by developing organisms. In the heat shock response, a whole battery of genes is turned off and another battery of genes turned on, in a matter of minutes, in response to some signal or stimulus. Embryonic development probably proceeds in such a fashion. A tissue might, for instance, make a decision to be anterior or posterior, dorsal or ventral, thoracic segment or abdominal segment, by switching on a series of related genes. Thus, the heat shock response is useful as a model system to study the mechanisms of coordinate gene regulation.
II. Procedure A – Polytene Chromosome Isolation and Staining Making good salivary gland chromosomes is almost an art. It will take some practice to get good, reproducible chromosomes. The stain (aceto orcein) can be administered for 5-30 minutes; the longer times will give darker staining. Squashing can be done with a thumb, a rubber stopper, or the eraser ball of a pencil. Student preparations are usually not squashed enough, so the chromosomes are in a tight end and are not splayed out enough for further analysis. Learning to recognize individual chromosomes and puffs is also a skill. With patience, care, and practice, each band and puff can be identified. It is not possible to become an instant expert. When the larva is subjected to environmental stress, its normal genes shut off and a new set is transcribed. The normal proteins stop being translated, and HSP proteins are made. A. Dissection of the salivary glands and polytene chromosome preparation 1. Place a few drops of insect Ringers or Saline in a well of a depression slide, watchglass, or small Petri plate. 2. Select a third instar larva from your bottle; they will be found near the bottom of the bottle, as they are just beginning to crawl up the sides. If the one you have chosen is sessile, it has already pupated and is of too late a stage to be used. Remove the larva with a pair of forceps or a dissecting needle and place in the well of your slide. 3. Under intermediate power of the dissecting microscope, place one forceps firmly across the rear half of the larva to prevent movement. Place the second forceps just behind the mouth hooks at the head region. The head is sharply pointed and the mouth hooks are black and so should be distinguishable from the rear of the larva. 4. Pull the head off the body by firmly pulling the two forceps apart. If you are successful, you should see the head with the two attached salivary glands trailing behind. The salivary glands can be recognized as two long, transparent, sausage-shaped bags with a characteristic translucent fat body along one side and occasionally capping each gland. Always keep the gland moist in Ringers solution.
B. Puffing pattern of salivary gland chromosomes 1. Transfer the glands immediately into a drop of 45% acetic acid placed on a microscope slide (frosted end up). Allow to remain approximately 30-45 seconds. Be sure not to let the glands dry out. 2. Transfer the glands to a small drop (2-3 mm in diameter) of aceto orcein stain in the center of the same slide. This should all be done under the dissecting scope to insure that the glands don't stick to the forceps. Leave them in the stain for 15-20 minutes (If the chromosomes aren't stained darkly enough, a longer incubation period may be necessary, last year 30 minutes worked well). Again, make sure the stain doesn't dry out during this time period. 3. Gently place a coverslip over this preparation. Put a piece of blotter paper over the coverslip and place a piece of parafilm over the blotter paper. 4. Place your thumb on the parafilm directly over the coverslip and press straight down quite firmly for about 20 seconds. You do not want the coverslip to move relative to the slide. Sometimes a #8 rubber stopper works well here. 5. Examine the slide under 10X to locate the squashes. Then, look under 40X to find a good preparation. Ideally, the chromosomes should be well spread and the bands stained darkly. Once you have found such a squash, place a drop of oil on top of the coverslip and examine under 100X. Now, try to identify the chromosomes based on the landmarks familiar to you.
D. Heat Shocked Larvae Once you have mastered the technique of squashing salivary glands, do a polytene preparation on larvae that have been heat shocked. They will be placed in the 37EC incubator about 30 minutes prior to when you will need them, and so the heat shock puffs should be of maximum size. Try to identify the heat shock puffs that were listed in the introduction. Note the regression of the other puffs normally found in this stage larvae (like the Chinese lanterns and the ballet skirt). This is indicative of a change in transcriptional activity due to this environmental insults.
II. Procedure B - Imaginal Disc Isolation (takes 2 days to observe effect) One of the many reorganizational processes that occur during the development of Drosophila is the evagination of imaginal discs into adult structures such as eyes, antennae, wings, legs, halters, and genitals. Imaginal discs are self-contained clusters of epithelial cells in insect larva that form early in embryonic development from small invaginations of ectoderm. As the embryo, and then the larva, grows, so will the imaginal discs. Although there are definite changes in imaginal disc size as the larva grows, the change in shape and conformation of the original pouch is most intriguing. Because growth occurs within a confined space, a fully developed larval imaginal disc resembles a flattened coil or collapsible camping cup. There are a total of nine pairs of imaginal discs (labrum, labial, antennal, eye, wing, three legs, and halters) plus a single gonadal disc. The identity of these discs is established by the anterior-posterior pattern of homeobox genes. (HOM complex). Imaginal discs are quite small during the 1st- and 2nd-instar stages, but begin to proliferate during the 3rd-instar stage. During pupation, the hormone ecdysone is secreted and initiates evagination of the imaginal discs. The centralmost portions of the discs will evaginate first and form the most distal part of the resulting structure. Once the discs have been exposed to ecdysone, they will telescope out into adult structures within 24 hours. Today you will dissect imaginal discs out of 3rd-instar Drosophila virilis larvae, expose discs to different concentrations of ecdysone, and monitor the results over 24 hours. You may also artificially evaginate some imaginal discs by exposing them to a proteolytic enzyme solution. Although you may have some difficulties in finding all imaginal discs, it is quite easy to locate the most anterior discs (especially the legs and eye-antenna). Drosophila is a new model system for us, and with all model systems there special techniques that may be mastered. Below are some common challenges with working with fly larvae:
Can’t find discs These are the most obvious near the brain, so if you can’t actually distinguish the discs, transfer the brain and surrounding tissues into the treatment solutions. Odds are that you will have transferred some discs as well. Didn’t transfer any of the discs you found Practice with bits of debris before you begin the experiment to get a feel for the microneedles. All parts of the transfer should be done under the dissecting microscope so you can see what you are really doing. Nothing telescoped You might not have had any discs in your treatment chamber. The enzyme might have become inactivated. Never make the solution up far in advance.
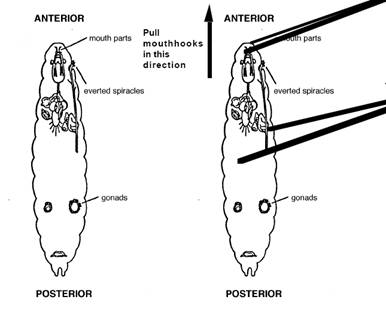
Location of imaginal discs in third instar Drosophila larvae and the first step on the dissection to isolate the discs.
1. Sterilize instruments in 70 % ethanol for 10 minutes 2. Prepare your four Petri dishes by filling each with 2 ml of serum-supplemented Schneider’s Insect Medium, then: a. Add 5 µl ecdysone solution to the first dish; b. Add 2 µl ecdysone solution the second dish; c. Add 1 µl ecdysone solution the third dish; d. Add 5 µl 95 % ethanol to your fourth dish as a control.
Be sure to label each dish! Don’t forget to add your name! 3. Obtain a 3rd-instar larva, place it on a clean slide, and rinse any debris off with Insect saline. 4. Transfer the larva to another sterile Petri dish and dissect out as many imaginal discs as you can. Grasp the head end of your larva just behind the mouth parts with one pair of forceps. Grasp the middle of the larva with the other pair and gently pull the body away from the head. The imaginal discs will be concentrated in the anterior third of the body, along the midline and attached to the tracheoles or gut. 5. Very gently transfer a few discs into each of the prepared solutions using the microneedles. 6. Sketch each disc and try to identify which discs you have. 7. Observe the discs every 30 minutes for 2 hours, looking for the beginning of eversion. 8. Allow the discs to incubate for 24 hours at room temperature. 9. Sketch and describe the discs. If you are not seeing many changes, you can treat the discs with trypsin which helps release the structures present in the discs, here is the protocol: a. Place 2 ml of the 1 % trypsin solution in a sterile Petri dish. b. Transfer several imaginal discs into the prepared dish. c. Observe the discs under a dissecting microscope for 60 minutes. Did the discs evaginate? e. Transfer an evaginated disc to a depression slide, view under the compound microscope and sketch.
ALL OBSERVATIONS SHOULD BE RECORDED IN YOUR LABORATORY NOTEBOOK. III. Data Analysis 1. Polytene chromosomes are found in nondividing cells only (for example, in salivary gland cells and intestine cells). Why not in actively dividing cells? 2. Why is it necessary to "fix" a specimen in acetic acid for microscopic observation? 3. Propose a likely reason why regions along the polytene chromosome (bands) stain more darkly than other spots (interbands). 4. Describe the location of heat shocked puffs that differ from normal chromosomes. 5. Did all the discs telescope out? 6. Were some discs more sensitive to the ecdysone? to the trypsin? 7. How do the ecdysone and trypsin treatments compare?
V. Materials 1. Wild-type Drosophila melanogaster (for polytene chromosomes) cultures can be obtained from most biological supply houses, such as Ward's Natural Science Establishment, Inc. (5100 West Henrietta Road, P. O. Box 92912, Rochester, New York 14692-9012) and Carolina Biological Supply Company (2700 York Road, Burlington, North Carolina 27215). Set up the cultures early and expand the colony to have one culture bottle per two students. Use only last-instar larvae; these are easy to identify because they crawl out of the medium and up the interior surfaces of the culture jar. Drosophila virilis 3rd-instar larvae (these will be crawling up the incubation chambers) are the best for imaginal disc dissection 2. Insect saline and aceto-orcein stain are prepared as follows:
Insect Saline 1.87 g NaCl 0.875 g KCl 0.078 g CaCl2
Aceto-orcein Stain 6 g orcein powder dissolved in a hot mixture of 150 ml glacial acetic acid 60 ml lactic acid 90 ml distilled water Filter twice. This stain will store indefinitely at 4EC. 3. A current genome map showing the identified bands and gene loci in Drosophila melanogaster chromosomes is available from Science, the journal of the American Association for the Advancement of Science (1333 H St., N.W., Washington, D.C. 20005). 4. Another source of genome maps is Lindsley, D. L., and Zimm, G. G. (1992). The Genome of Drosophila melanogaster. Academic Press, Inc. (1250 Sixth Avenue, San Diego, CA 92101). Eight polytene maps of C. B. Bridges, P. M. Bridges, and G. Lefevre are bound into this reference book and are also available separately as an eight-map packet for student use. Toll free ordering available: 1-800-321-5068 or FAX 1-800-336-7377. 5. Pasture pipettes and bulbs and coverslips 6. Slides, coverslips, and depression slides, 70% ethanol squirt bottles 7. Sterile Insect pins and watchmakers forceps any microsurgery tools you have made 8. Petri dishes with moist filter paper bottoms (to hold slides while they stain) 9. 45% acetic acid solution 10. Dissecting scopes with lights and microscopes with oil immersion lenses, lens oil and lens paper 11. TV monitor connected to video camera on microscope 12. Small paintbrush (for removing larvae from the incubation chamber) 13. Sterile 34-mm Petri dishes or a multiwall tissue culture 14. Micropipettors (1-5µl and 200-1000µl) 15. Sterile 2-5 ml glass pipettes 16. 70% and 95% squirt bottles of ethanol 17. Schneider’s Insect Medium supplemented with 10 % fetal calf serum (containing growth factors) 18. Ecdysone solution (1 mg ecdysone/1 ml 95 % ethanol) 19. 1 % trypsin solution in Insect saline
V. References 1. Biroc, S. L. (1986). Developmental Biology: A Laboratory Manual with Readings, pp. 154-169. New York: Macmillan Publishing Co. 2. Bridges, C. B. (1935). Salivary chromosome maps with a key to the banding of the chromosomes of Drosophila melanogaster. J. Hered. 26, 60 and supplement. 3. Lefevre, G., Jr. (1976). A photographic representation and interpretation of the polytene chromosome of Drosophila melanogaster salivary gland. In The Genetics and Biology of Drosophila, Vol. 1a, pp. 31-66 (M. Ashburner and E. Novitski, eds.). London: Academic Press. 4. Lindsley, D. L. and Zimm, G. G. (1992). The Genome of Drosophila melanogaster. San Diego: Academic Press. 5. Ahern, H. (1992). Introduction to Experimental Cell Biology. WMC Brown Publishers 6. Merrian, RW. (1988). Experiments in Animal Development. Sinauer Associates Inc. Publishers. 7. Http://1vm.harvard.edu.7081/images/1k/contributed-images/csomes-weeks-etal/ 8. Gibbs, MA (2003). Practical guide to developmental biology. Oxford
|