![]()
|
|
|
LABORATORY TEN PLANT STRUCTURE AND EVOLUTION OVERVIEW: This laboratory, like the remaining one on animal diversity, is designed to emphasize observation of material rather than activity-like exercises. Therefore, it is of paramount importance that you read this material before lab and refer to it as you examine the wide range of slides, specimens, and demonstrations available in the laboratory. The introductory material combines very specific information about three different life histories, along with an evolutionary account of how plants adapted to life on land. You will use both compound and dissection microscopes to study ferns in detail. You will observe demonstrations concerning the major groups of the Kingdom Plantae, focusing on their specific adaptations for life on land. Think about how the structures function as solutions to environmental challenges.
OBJECTIVES OF LABORATORY: • To learn the major characteristics of plants. • To become familiar with basic plant life history and its appropriate terminology. • To become familiar with the life history of the fern as a representative plant. • To recognize basic structures of plant types. • To become familiar with adaptations in plants that make terrestrial existence possible. • To practice observation skills with the compound and dissection microscopes.
INTRODUCTION: While all life began in the oceans, today the surface of the land is also rich with life. Land animals could not exist without the primary productivity of land plants. Yet the fossil record shows that for hundreds of millions of years after the origin of life, terrestrial plants were absent. The invasion of land by plants may have begun as recently as 450 million years ago. Terrestrial plants profoundly modified the land environment, adding organic matter to soils as they decayed, reducing erosion by binding sediments with their roots, speeding up the weathering of rocks by releasing organic acids, and providing a food source for animals, bacteria and fungi. Undoubtedly, they also changed the atmosphere, removing carbon dioxide and releasing quantities of oxygen. Compared to marine algae, or seaweeds, most land plants have complex structure. Seaweeds, despite a variety of forms, do not have roots, stems or leaves, only differently shaped regions of their bodies that serve similar functions. Land plants, on the other hand, have these three basic plant organs, which anchor them and obtain water (roots), act as photosynthetic platforms and gas exchangers (leaves) and conduct material both ways between roots and leaves (stems). Their cells are more strongly differentiated, and there are more kinds of them. In particular, they have conducting tissues that serve to move water and nutrients around their bodies. These range from very simple tubes in mosses to highly evolved xylem and phloem in vascular plants. Land plants also have cuticle, a secreted layer on the outside of their bodies to help limit water loss. The cuticle also limits gas exchange, so it must be interrupted here and there with pores called stomates that allow plants to "breathe." Fundamental changes were also needed in the plant life cycle for plants to survive on land. Plants have a form of life cycle known as alternation of generations (Fig. 10.1). Sexually reproducing, haploid individuals (gametophytes) alternate generation for generation with asexually reproducing, diploid individuals (sporophytes). Meiosis takes place in the asexual individuals, producing haploid spores. This is in contrast to the process of sexual reproduction in animals, where meiosis gives rise to gametes. Plant gametes (eggs and sperm) are produced through mitosis, which takes place in a plant body that is already haploid.





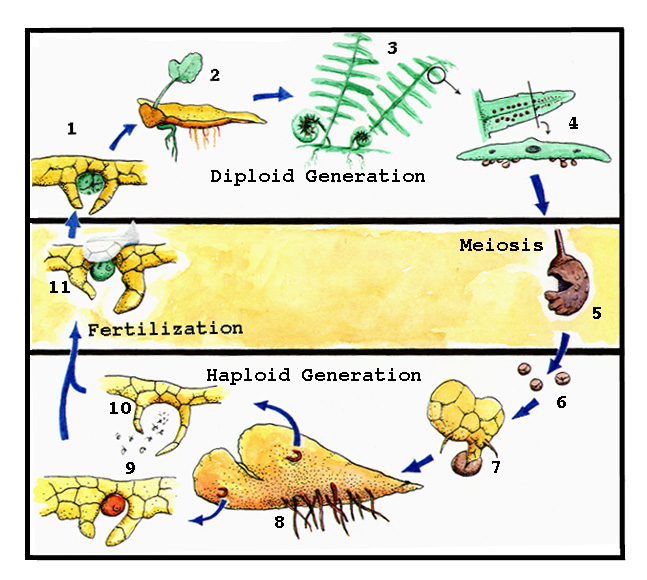
Life Cycle of Plants: Alternation of Generations FIGURE 10.1
Probably the first step in adapting to land was to produce spores that could stand being dried out. A number of algae still living today do this. The advantage is that the spores can become part of the dust and be carried to new habitats by the wind, or can wait in the dried mud until water again fills their pond. Naturally, because they are enclosed in a hard coat, these spores could not swim. The part of the sporophyte that produced them might have been adapted to extend above the water surface; it would need a cuticle to prevent drying out. However, the gametophyte generation remained unchanged; perhaps just one copy of each gene was not enough to allow for the development of new kinds of structures. The gametes became differentiated so that sperms and eggs evolved. The sperms are small and motile, while the eggs are larger and are retained in the body of the female gametophyte plant. Sexual fusion (fertilization) can only occur if there is water available in which the sperm may swim to the egg. The most primitive land plants alive today are the liverworts, of which Marchantia is an example (see Fig. 10.3). The most conspicuous part of the life cycle is the gametophyte, which looks something like a small seaweed tightly pressed to damp soil. Without a cuticle, and with breathing pores that cannot be closed, the gametophyte can easily dry out and die. Marchantia is limited to wet or damp habitats. Periodically, male plants produce umbrella-like structures that carry antheridia, sperm-producing organs, and female plants produce palm-tree-like structures that carry archegonia, where single eggs arise. During a rain or on a dewy morning, the sperm have to swim all the way to a female plant and find the opening to an archegonium. They are guided by chemical signals. Most of them don't make it, but huge numbers are released, so nearly every egg does get fertilized. The resulting zygotes stay on the female gametophyte and develop into small, capsule-like sporophytes. The sporophytes do not carry on photosynthesis and are actually parasites of their female parent. Meiosis occurs inside the capsule, and eventually spores are released. The spores germinate on wet soil to produce another generation of gametophytes. Because the gametophytes can't resist drying out and because the sperm have to swim through water to reach the eggs, liverworts can't be regarded as very well adapted to life on land. Mosses are more common relatives of liverworts and share many features of their life cycle. There are even desert-dwelling mosses, but they still can't reproduce sexually unless a film of water is available in which the sperm can swim to the egg.
Life Cycle of LiverwortFigure 10.3
The liverworts and mosses were probably not the ancestors of more highly evolved land plants, though their characteristics suggest to us what those ancestors might have been like. Probably they did not have such small, parasitic sporophytes. The two generations may have been similar in appearance and size. Many groups of such plants are known from fossils found in the Silurian and Devonian Periods of the earth's history, beginning about 420 million years ago. Most of them became extinct without leaving any descendants, but one group evolved into ferns. The fern life cycle is rather similar to the liverwort, but the size relations of the sporophyte and gametophyte generations are reversed (see Fig. 10.4). The gametophytes, instead of dominating the life cycle, are small, very simple, and usually short-lived; most people have never seen them. The sporophytes, on the other hand, are the large, obvious plants we call "ferns." Some species reach the dimension of small trees and most live for many years. Annually (continuously in the wet tropics) they produce dot-like masses of spore capsules (sporangia) on the lower surfaces of their fronds, or leaves. These masses are called sori. With thousands of sori, a single fern can release billions of haploid spores. Most of these are wasted. A few may land on damp soil or rock and germinate into gametophytes, called prothalli. Each prothallus, usually smaller than the nail of your little finger, produces several archegonia and antheridia. Again, the motile sperm must swim to the eggs in a film of water. The resulting zygote slowly grows into a new sporophyte as the gametophyte disintegrates. We’ll view a short film that shows all these events. Fern sporophytes are advanced land plants. They have a thick cuticle with pores that can open and close, and very efficient systems of tubular cells to conduct water up from the roots and food down from the leaves. They can send out long thin stems with buds on them and rapidly spread to cover large areas. With the appearance of such sporophytes, perhaps as early as 400 million years ago, most of the physical adaptations of plants to land were complete. But they remained reproductively limited because the gametophyte requires very damp conditions that last at least long enough for it to produce gametes and get the new sporophyte started. Relying on vast numbers of tiny spores also seems inefficient. In ecological terms, these factors cause ferns to be regarded as opportunistic, or r-selected, organisms, colonizers of unstable, transient habitats. About 375 million years ago, late in the Devonian Period, a group of ferns arose that modified this reproductive strategy. They have been called Seed Ferns. Their sporophytes were very much like those of other ferns, but their spores were never released from their sporangia. Instead, they were retained and germinated into tiny, parasitic gametophytes right on the fern fronds. The eggs of the female gametophytes were also fertilized there, and the new sporophytes started development while attached to their "parents," which were in turn attached to their "grandparents." Each female gametophyte with its enclosed sporophyte embryo was packed with stored food material by the "grandparent" sporophyte and covered with a hard outer casing. In other words, the seed had evolved! The problem of the wasteful production of billions of spores had been solved. The number of spores could be limited because each spore germinated into a protected gametophyte. Each seed consisted of an embryo sporophyte with stored food, which could give it a boost of rapid early growth. Such plants were well adapted to succeed in habitats where competition was high and where populations were always near carrying capacity. They were K-selected, or equilibrium, organisms.
Life Cycle of a FernFigure 10.4 1. Zygote (2n) 6. Spores 2. Immature Sporophyte 7. Young Gametophyte 3. Mature Sporophyte 8. Mature Gametophyte 4. Leaf with Sori 9. Archegonium 5. Sporangium 10. Antheridium 11. Fertilization From among these seed ferns, true seed plants emerged. The earliest of these are called Gymnosperms, and they are still around in the form of pines, spruces, hemlocks, and a few more obscure sorts, such as gingkos. The seed ferns that were the ancestors of gymnosperms had added a new wrinkle. Instead of having motile sperm that had to swim to the egg, they reduced the entire male gametophyte to a tiny structure of only a few cells, and enclosed it in a hard casing. These tiny objects could easily be carried by the wind and some of them would land on the female gametophytes. The minute male gametophytes, pollen grains, can produce only one sperm, but that is all that is needed to fertilize the egg. Each of the countless pine pollen grains that dust our cars and houses in spring is actually a tiny, independent plant! The pollen of pines is produced in tight clusters of modified leaves that make up a cone, or strobilus. The male cones release their pollen and disintegrate. The female cones are what we know as pinecones. They are woody and can be very large, as much as two feet long. Each cone scale is a modified leaf and bears a sporangium at its base. The megasporangium is covered in one or two layers of cells, and produces a small number of female megaspores. One of these germinates and slowly grows into a female gametophyte, which, by mitosis, gives rise to eggs. The whole thing, outer cell layers, megasporangium, female gametophyte and egg, is called an ovule. Wind carries pollen to the sticky receptive surface of the ovule. Recent experiments in wind tunnels show that pinecones are exquisitely designed to conduct air currents within, and pine pollen is produced so abundantly that most ovules get some. The pollen grain cracks open, and a tube emerges that penetrates the ovular tissues. It takes a year or longer for this tube to grow to the egg, at which point the single sperm travels down it and fertilizes the egg. The ovule is then converted into a seed as the embryo grows and food is stored in the tissues of the old female gametophyte. Finally, the cone opens and the seeds are released. The life cycle of the pine therefore appears to skip the gametophyte generation entirely. Only those lucky enough to have taken biology know that this is not true and that the gametophyte generation still exists in the form of pollen grains and ovules. Finally, probably around 250 million years ago in the Triassic Period, land plant evolution culminated in the Angiosperms, or flowering plants. It is not at all clear which of many kinds of gymnosperms alive in the Triassic evolved into flowering plants. However, the appearance and rise of certain kinds of animals may have triggered this evolutionary change. Flowering plants produce very small amounts of pollen compared to gymnosperms; a saving possible because they rely on insects to carry the pollen to another plant of their species. The insects are attracted by modified clusters of leaves called flowers, and by rewards of food produced there. In addition, the angiosperms cover their seeds with a fleshy fruit, relished by vertebrates or other large, plant-eating animals. While the fruit is digestible, the seeds it contains pass unharmed through the digestive system, emerging many hours later when the animal is perhaps far away. In this fashion, the seeds can be widely distributed. Testimony to the effectiveness of this method can be found in the millions of tomato plants that sprout at the margins of sewage treatment lagoons! The angiosperms have also carried the reduction of the gametophyte generation even further, but the technicalities of this life cycle are not really needed to complete our story of the invasion of the land. With the evolution of flowers and fruit, angiosperms have become so diverse and abundant that they now dominate the plant life of nearly all terrestrial biomes. Only the gymnosperms can challenge their dominance in some special habitats, such as the taiga or the southern pine forests. In the coal swamps of the Carboniferous, 300 million years ago, ferns, seed ferns, gymnosperms, lycopods (ground pines) and sphenopsids (horsetails) all reached tree stature. The success of the angiosperms has produced a much more monotonous, but arguably more beautiful, world.
Life Cycle of a PineFigure 10.5
LABORATORY INSTRUCTIONS: To begin our study of the life cycle of ferns, we will view a video of this interesting organism that exhibits clear alternation of generations in its life history. The video has commentary, but your instructor will add to your study specific points about the ferns' adaptation to land habitation and also point out when ferns need to be in or near water to continue the life cycle. After the video, begin your study with the mature fern plant. Make drawings and answer the questions on the report sheet included here. The drawings that are required for the report sheet are underlined in the directions here.
The Fern Life History 1. At your lab bench locate a mature fern in a pot. The leaves of the fern are called fronds. The young leaves are referred to as "Fiddleheads" because they are coiled when they are first produced. The individual leaflets are called pinnae. This familiar form of the fern is the sporophyte generation, the asexual generation. It is on the underside of the mature leaves that meiosis occurs to produce spores.
2. Notice that the stem and roots of the mature fern are beneath the soil in the pot. The fern stem is called a rhizome an underground stem. Examine the prepared slide of the cross-section of a fern rhizome. Notice the round or oval areas that are vascular bundles. The cells of the vascular bundles are: xylem, the hollow red-stained cells, that conduct water up to the fronds from the roots, and the less obvious phloem cells that move the products of photosynthesis up and down the plant as needed. Draw and label the cross- section of a vascular bundle of the fern rhizome showing some cellular detail. In what ways is vascular tissue an adaptation for life on land? Observe the uncovered rhizome with its roots on demonstration in the laboratory. The sporophyte can continue to grow asexually by extending the length of the horizontal stem and sending out roots and sending up new fronds. |